 聯系我們
聯系我們免疫系統爲入侵的病原體提供了(le)關鍵的防禦,面對(duì)對(duì)小(xiǎo)分子藥物的耐藥性上(shàng)升和(hé)新抗菌劑管道(dào)的萎縮,這(zhè)些(xiē)病原體變得越來(lái)越重要。保護性抗體是任何免疫反應的關鍵組成部分,因爲它們可以中和(hé)分泌的毒素,阻斷與宿主細胞的相互作(zuò)用(yòng),并招募免疫系統防禦。這(zhè)些(xiē)抗體可以通過自(zì)然感染或疫苗接種引起,也(yě)可以作(zuò)爲純化的蛋白(bái)質被動給藥。
目前單克隆抗體藥物已用(yòng)于五種傳染病:呼吸道(dào)合胞病毒(RSV)、炭疽、複發性艱難梭菌、SARS-CoV-2和(hé)埃博拉病毒。事(shì)實證明(míng),開(kāi)發針對(duì)更複雜(zá)的病原體的有效疫苗和(hé)抗體藥物具有挑戰性,因爲自(zì)然免疫力通常不足以防止再感染。這(zhè)些(xiē)病原體編碼多種蛋白(bái)質,其表達由複雜(zá)的調控途徑協調,在不同的疾病狀态下(xià)表達的抗原很(hěn)少。功能(néng)冗餘毒力因子和(hé)菌株變異的表達會(huì)使靶标選擇進一步複雜(zá)化。此外(wài),許多病原體已經進化出多種免疫逃避策略,在某些(xiē)情況下(xià),可以在吞噬作(zuò)用(yòng)後避免消除。即使是基因相對(duì)較少的病原體,如流感和(hé)SARS-CoV-2病毒,也(yě)設法迅速逃避針對(duì)少數靶向蛋白(bái)質的免疫反應,這(zhè)強調了(le)對(duì)更複雜(zá)的抗體的需求。因此,在上(shàng)篇介紹針對(duì)病原體保護脆弱的抗原表位進行重組抗體的設計(jì)和(hé)開(kāi)發策略之後,我們今天總結下(xià)靶向破壞抗體Fc功能(néng)的病原體進行重組抗體的設計(jì)和(hé)開(kāi)發策略。
盡管有許多努力集中在開(kāi)發中和(hé)或封閉抗體,但(dàn)Fc效應功能(néng)在保護作(zuò)用(yòng)中的作(zuò)用(yòng)越來(lái)越得到(dào)認識。這(zhè)些(xiē)可由人IgG1和(hé)IgG3同種型介導,後者與宿主Fc受體結合,通過自(zì)然殺傷(NK)細胞、中性粒細胞和(hé)巨噬細胞的抗體依賴性細胞吞噬作(zuò)用(yòng)以及補體依賴性細胞毒性介導ADCC。這(zhè)些(xiē)反應依賴于Fc與補體成分C1q和(hé)經典激活性Fc受體CD16A和(hé)CD32A的相互作(zuò)用(yòng),而這(zhè)兩種受體高(gāo)度依賴于靶細胞表面的抗原密度和(hé)表位可及性。雖然Fc相互作(zuò)用(yòng)可導緻抗體依賴的增強型疾病,尤其是在登革熱感染中,但(dàn)它們通常代表了(le)适應性免疫應答(dá)的強大(dà)力量,可以補充中和(hé)抗體。不幸的是,這(zhè)也(yě)爲病原體提供了(le)選擇壓力,使其通過降解或隔離抗體來(lái)逃避這(zhè)些(xiē)功能(néng)。針對(duì)抗裂解、阻斷蛋白(bái)酶活性或抗抗體隔離而設計(jì)的抗體爲消除這(zhè)些(xiē)免疫逃逸策略提供了(le)機會(huì)。
抗緻病性蛋白(bái)酶活性的抗體
木(mù)瓜蛋白(bái)酶和(hé)胃蛋白(bái)酶等酶通常通過分别在上(shàng)鉸鏈區(qū)和(hé)下(xià)鉸鏈區(qū)切割序列來(lái)産生Fab和(hé)F(ab ')2抗體片段。幾種與腫瘤侵襲和(hé)炎症相關的人基質金(jīn)屬蛋白(bái)酶(MMPs),如MMP-3和(hé)MMP-7,也(yě)可以切割免疫球蛋白(bái),這(zhè)表明(míng)這(zhè)些(xiē)特征在系統發育過程中是常見的。作(zuò)爲免疫逃逸的一部分,細菌可以分泌具有同源功能(néng)的酶。例如,鏈球菌熱原性外(wài)毒素B (SpeB)、免疫球蛋白(bái)g降解酶(IdeS)和(hé)内糖苷酶(enos)是由化膿性鏈球菌和(hé)裂解抗體分泌。SpeB對(duì)免疫球蛋白(bái)有廣泛的特異性,ide對(duì)IgG鉸鏈有特異性,而EndoS會(huì)去除與Fc受體結合所需的N297殘基上(shàng)的糖。類似地,金(jīn)黃色葡萄球菌産生谷氨酰内肽酶V8 (GluV8),該酶在上(shàng)鉸鏈區(qū)切割免疫球蛋白(bái),而銅綠假單胞菌分泌大(dà)量的彈性蛋白(bái)酶B (LasB),該酶也(yě)可能(néng)在鉸鏈區(qū)切割免疫球蛋白(bái)。
雖然生物技術的應用(yòng)是有效的,但(dàn)抗體裂解可能(néng)不利于保護性免疫反應。事(shì)實上(shàng),添加蛋白(bái)酶抑制劑降低(dī)了(le)抗體切割,同時(shí)增加了(le)C3b補體沉積和(hé)中性粒細胞對(duì)金(jīn)黃色葡萄球菌的吞噬。大(dà)多數IGg特異性蛋白(bái)酶采用(yòng)兩步過程:一個重鏈鉸鏈被裂解,産生一個中間産物,然後第二個鉸鏈被緩慢裂解,産生Fc和(hé)Fab或F(ab ')2片段。單獨切割的IgG保留抗原結合活性,但(dàn)不再促進效應功能(néng)。在許多個體中觀察到(dào)與裂解的IgG1結合的自(zì)身抗體,這(zhè)表明(míng)這(zhè)一中間産物具有免疫原性和(hé)生理(lǐ)學意義。裂解抗體在細菌表面的累積會(huì)抑制完整抗體與抗原的接觸,從(cóng)而減少Fc效應功能(néng)的募集。因此,即使是不完全的抗體裂解也(yě)能(néng)有效地逃避免疫反應。
爲了(le)恢複因抗原結合抗體和(hé)裂解抗體而喪失的Fc效應細胞功能(néng),研究者探索了(le)使用(yòng)可識别裂解産生的新表位的抗體(Figure 2)。這(zhè)些(xiē)完整的抗體可以重建靶細胞和(hé)吞噬細胞上(shàng)Fc受體之間中斷的聯系。Jordan等通過在兔感染模型中使用(yòng)與GluV8快(kuài)速裂解的金(jīn)黃色葡萄球菌表面結合的抗體來(lái)評估這(zhè)種方法。作(zuò)者表明(míng),使用(yòng)類似于抗體鉸鏈序列的肽進行免疫可引起特異性識别gluv8裂解抗體的強抗體反應。與未免疫組相比,用(yòng)鉸鏈樣肽免疫的兔的金(jīn)黃色葡萄球菌定植水(shuǐ)平降低(dī),并且在體外(wài)試驗中,這(zhè)些(xiē)動物的血清恢複了(le)補體依賴性細胞毒性。
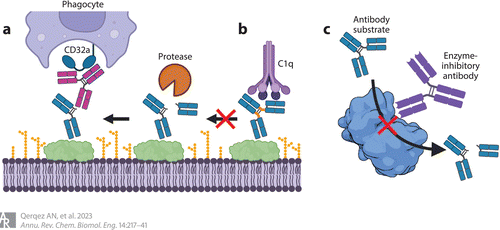
Figure 2 Antibodies that resist pathogenic protease activities to sustain recognition by host immune proteins, such as the C1q component of complement and CD32a Fc receptor found on phagocytes. (a) Antibodies recognizing hinge epitopes exposed after cleavage restore Fc functions to cleaved antibodies. (b) Antibodies with engineered hinge regions no longer serve as suitable substrates for pathogenic proteases. (c) Antibodies that block the activity of pathogenic proteases by directly blocking access to the active site (shown) or noncompetitive allosteric mechanisms (not shown) can protect antibody functions. Shown is the structure of LasB (PDB 3DBK). Adapted from images created with BioRender.
構建抗裂解抗體是對(duì)抗蛋白(bái)水(shuǐ)解降解的另一種策略。雖然IgG1是人體内含量最多的同種型,也(yě)是藥物性抗體最常見的同種型,但(dàn)IgG2通常對(duì)蛋白(bái)水(shuǐ)解性裂解更有抗性。然而,與IgG1相比,IgG2與CD16a和(hé)C1q的結合更弱,并相應地介導減少Fc依賴性殺傷和(hé)補體沉積。将IgG2的彈性與IgG1的功能(néng)相結合的努力導緻了(le)嵌合體,其中IgG2的下(xià)鉸鏈和(hé)CH2序列取代了(le)IgG1的。然而,抗體也(yě)通過在保守的下(xià)端鉸鏈與經典Fc受體結合,這(zhè)一區(qū)域是許多蛋白(bái)酶的底物。因此,增加抗體抗裂解能(néng)力的突變也(yě)對(duì)Fc受體結合和(hé)免疫效應功能(néng)的募集産生負面影響。
爲了(le)解決對(duì)抗體蛋白(bái)酶抗性的需求,同時(shí)保留效應物的功能(néng),我們産生了(le)IgG1/IgG2混合抗體。Kinder等首先通過引入E233P/L234V/L235A替換和(hé)删除Gly236,将IgG2的下(xià)鉸鏈引入到(dào)IgG1中。正如預期的那樣,在體外(wài)試驗中,這(zhè)種結構失去了(le)促進補體殺傷和(hé)吞噬的能(néng)力。之前确定的選擇性激活補體殺傷或調理(lǐ)吞噬作(zuò)用(yòng)的突變随後被引入該嵌合體。變異體2h-DE (S239D/I332E)恢複了(le)Fc受體結合,而變異體2h-AA (K326A/E333A)恢複了(le)補體殺傷。結合這(zhè)些(xiē)突變産生2h-DAA (239D/K326A/E333A)和(hé)2h-AEA (K326A/I332E/E333A)恢複了(le)補體殺傷和(hé)調理(lǐ)吞噬活性,同時(shí)保持了(le)對(duì)多種蛋白(bái)酶(化膿性鏈球菌IdeS、金(jīn)黃色葡萄球菌GluV8、MMP-3和(hé)MMP-12)切割的抗性(圖2b)。蛋白(bái)酶耐藥Fc結構域的加入可能(néng)支持藥物性抗體的開(kāi)發,以用(yòng)于細菌感染(細菌感染的庫中包括抗體降解蛋白(bái)酶)。
用(yòng)中和(hé)抗體直接靶向蛋白(bái)酶是阻斷蛋白(bái)水(shuǐ)解活性的一種替代方法,具有阻斷所有底物裂解的優點。這(zhè)種方法最初用(yòng)于腫瘤相關的MMPs,從(cóng)而鑒定出Kivalues ~ 5 nM(78)作(zuò)爲競争性抑制劑的抗體。它擴展到(dào)銅綠假單胞菌假溶素(LasB)蛋白(bái)酶,該酶通過切割彈性蛋白(bái)、膠原、IgA、IgG和(hé)補體蛋白(bái)來(lái)支持早期感染
抗蛋白(bái)水(shuǐ)解裂解的抗體可能(néng)提高(gāo)其保護能(néng)力,并且可以通過識别抗降解的Fc取代來(lái)實現(xiàn)。重組抗體工(gōng)程的一個吸引人的特點是,一旦開(kāi)發出合适的工(gōng)程Fc,它可以與結合任何抗原的Fab臂結合。或者,可以爲每一種感興趣的蛋白(bái)酶開(kāi)發酶抑制抗體。如果得到(dào)單克隆抗體研究的支持,經過适當工(gōng)程改造的蛋白(bái)酶免疫原可增強天然抗體和(hé)疫苗誘導的多克隆抗體産生的保護作(zuò)用(yòng)。
阻斷細菌fc結合蛋白(bái)質類的抗體
各種細菌病原體表達與保守抗體序列結合的毒力因子。這(zhè)些(xiē)捕獲的抗體可以屏蔽病原體,阻斷免疫對(duì)其他(tā)表面抗原的接觸,拮抗Fc效應功能(néng),甚至改變免疫細胞信号傳導。雖然多種生物分泌抗體結合蛋白(bái),包括金(jīn)黃色葡萄球菌第二免疫球蛋白(bái)結合蛋白(bái)(Sbi)、化膿性葡萄球菌蛋白(bái)G和(hé)M以及大(dà)胃鏈球菌蛋白(bái)L,但(dàn)研究最多的是金(jīn)黃色葡萄球菌蛋白(bái)A。蛋白(bái)A在所有定植株中均有表達,由5個同源結構域組成,每個同源結構域均能(néng)與除IgG3以外(wài)的所有人類IgG同型和(hé)除IgG1以外(wài)的所有小(xiǎo)鼠IgG同型的Fc高(gāo)親和(hé)力結合(KD值爲2.6-14 nM)。
當蛋白(bái)A通過c端錨定附着于細菌細胞壁時(shí),可阻斷抗葡萄球菌抗體對(duì)表面抗原的接觸。它還通過幹擾IgG六聚體形成和(hé)通過空(kōng)間阻斷Fc與低(dī)親和(hé)力Fc受體的相互作(zuò)用(yòng)(Figure 5)來(lái)防止抗體依賴性補體激活。
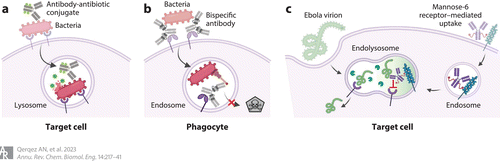
Figure 5 Antibodies that target intracellular pathogens. (a) Antibody–antibiotic conjugates bind bacterial surface antigens and are internalized with the bacteria by natural or phagocytic mechanisms. Once internalized, the antibiotic is released by resident enzymes to kill co-localized bacteria. (b) Bispecific antibody MEDI3902 uses one binding site to bind the Psl surface antigen on Pseudomonas aeruginosa and mediate phagocytosis. After translocation to the endosome, the second antibody-binding site blocks type III secretion to support endosome acidification and bacterial killing. (c) The mannose-6-phosphate receptor can mediate antibody transfer to an endolysosome that may already contain Ebola virions. Once co-localized, the antibody can block Ebola–receptor interactions to prevent viral escape into the cellular cytosol. Adapted from images created with BioRender.
此外(wài),一些(xiē)蛋白(bái)A從(cóng)細菌表面釋放(fàng),可與vh3型b細胞受體或抗體Fc結構域結合。B細胞受體交聯激活B細胞,導緻克隆擴增和(hé)凋亡塌陷,從(cóng)而限制抗葡萄球菌應答(dá)(Figure 3b)。與遊離抗體的Fc結構域結合可以在細胞循環過程中阻斷與新生兒Fc受體(FcRn)的相互作(zuò)用(yòng),從(cóng)而靶向抗體進行溶酶體降解(Figure 3c)。
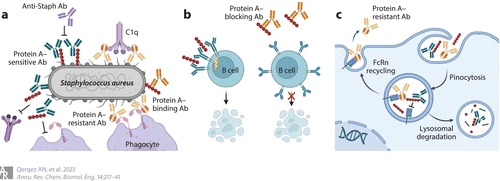
Figure 3 Antibodies that resist capture by Fc-binding protein A. Staphylococcus aureus protein A disrupts antibody responses in multiple ways but can be restored by Fc domains with reduced protein A affinity (most IgG3 allotypes or engineered IgG1 domains) or antibodies that bind protein A to block Fc capture. (a) Membrane-bound protein A can block antibody Fc binding to the low-affinity Fc receptors CD16b and CD32a, block the Fc hexamerization required for efficient recruitment of C1q to activate complement, and shield the bacterial surface from recognition by antibody Fab domains. Secreted protein A can (b) crosslink VH3 domains to trigger B cell receptor activation and apoptotic collapse and (c) block antibody recycling by Fc/FcRn binding to reduce antibody half-life. Unmodified antibodies are shown in green, Fc engineered antibodies are shown in yellow, and antiprotein A antibodies are shown in orange. Abbreviations: Ab, antibody; FcRn, neonatal Fc receptor. Adapted from images created with BioRender.
已經探索了(le)用(yòng)單克隆抗體的抗原結合抗原表位直接靶向蛋白(bái)A,以阻斷其Fc結合活性,支持細菌的調理(lǐ)吞噬作(zuò)用(yòng)。抗體514G3與在有血清抗體的情況下(xià)細菌表面可接近的蛋白(bái)a表位結合。在小(xiǎo)鼠模型中,在耐甲氧西林(lín)金(jīn)黃色葡萄球菌緻死性攻擊前給藥時(shí),該抗體顯示出保護作(zuò)用(yòng),而較低(dī)的抗體劑量與萬古黴素抗生素治療顯示出協同作(zuò)用(yòng)。
抗體3F6與蛋白(bái)A和(hé)相關蛋白(bái)Sbi結合,阻斷Fc捕獲和(hé)B細胞交聯效應(Figure3b)。用(yòng)3F6作(zuò)爲小(xiǎo)鼠IgG2a進行被動免疫可保護新生小(xiǎo)鼠免受血流感染,并使其血清中針對(duì)其他(tā)金(jīn)黃色葡萄球菌抗原的IgG滴度較高(gāo)。
人們一直在努力設計(jì)能(néng)引起強512G3和(hé)3f6樣蛋白(bái)A應答(dá)的抗原,目的是降低(dī)蛋白(bái)A對(duì)Fc的親和(hé)力。與野生型蛋白(bái)A不同,一個名爲SpAKKAA的初始解毒變異體,SpAKKAA免疫可誘導産生高(gāo)滴度抗體,從(cóng)而促進調理(lǐ)吞噬細胞殺傷,并保護小(xiǎo)鼠免受金(jīn)黃色葡萄球菌血流感染。然而,SpAKKAA持續引起動物應激,提示其保留了(le)部分B細胞超抗原活性。SpAKKAA等免疫原能(néng)更好(hǎo)地誘導抗體幹擾蛋白(bái)A的功能(néng),因爲即使在Fc存在的情況下(xià),關鍵表位仍然暴露。這(zhè)種方法爲設計(jì)其他(tā)Fc結合蛋白(bái)的免疫原提供了(le)希望。
抗Fc結合蛋白(bái)捕獲的抗體
爲了(le)支持開(kāi)發用(yòng)于疾病的單克隆抗體,可以對(duì)抗體Fc結構域進行改造,以降低(dī)對(duì)緻病性Fc結合蛋白(bái)的親和(hé)力。先前的研究表明(míng),由于聚焦化或氨基酸序列的改變,抗體對(duì)活化的CD16a和(hé)CD32a宿主受體具有更高(gāo)的親和(hé)力,從(cóng)而增強了(le)與癌症相關的效應功能(néng)。然而,這(zhè)些(xiē)和(hé)相關的Fc變化對(duì)感染性疾病反應的潛在影響還不太清楚。
爲了(le)确定抗蛋白(bái)A捕獲的抗體是否介導了(le)更強的金(jīn)黃色葡萄球菌吞噬作(zuò)用(yòng),我們選擇了(le)一種與豐富的表面糖共聚物壁磷壁酸結合的抗體。兩個降低(dī)蛋白(bái)A結合親和(hé)力的殘基改變(H435R和(hé)Y436F)被引入人IgG1 Fc結構域。類似Fc結合蛋白(bái)由疱疹病毒表達,當這(zhè)些(xiē)病毒Fc受體在感染細胞表面表達時(shí),它們可以捕獲與鄰近病毒糖蛋白(bái)結合的抗體Fc結構域。
這(zhè)種拮抗作(zuò)用(yòng)可抑制Fc與免疫細胞上(shàng)的宿主Fc受體的結合和(hé)信号傳導,并可導緻抗體内化,用(yòng)于溶酶體降解或重新包裝,使抗體覆蓋病毒粒子。同樣,當使用(yòng)缺乏病毒Fc受體的病毒株感染成纖維細胞時(shí),結合巨細胞病毒蛋白(bái)的抗體結合人Fc受體,并更有效地誘導ADCC。這(zhè)表明(míng),Fc結構域可以被設計(jì)來(lái)增強非中和(hé)性抗疱疹病毒應答(dá)(如ADCC)的效力。
防止病原體隔離抗體可以使宿主免疫系統有效地應對(duì)感染。在金(jīn)黃色葡萄球菌中,抗體514G3阻斷了(le)蛋白(bái)A與任何抗體Fc的結合,而使用(yòng)IgG3 Fc可防止蛋白(bái)A破壞治療性抗體活性。這(zhè)些(xiē)報(bào)告爲靶向蛋白(bái)A的抗體提供了(le)概念驗證,可能(néng)擴展到(dào)其他(tā)Fc結合蛋白(bái)。結合重組抗體技術的進步,我們認爲利用(yòng)抗體逃逸途徑的知(zhī)識開(kāi)發設計(jì)Fc結構域可介導針對(duì)特定病原體的有效效應功能(néng)。
獻")








