 聯系我們
聯系我們自(zì)1965年發現(xiàn)B細胞以來(lái)(Cooper et al. 1965),我們對(duì)B細胞的發育、成熟和(hé)功能(néng)的認識有了(le)巨大(dà)的進步。B細胞通過抗原呈遞、細胞因子分泌和(hé)自(zì)身抗體的産生,是多種自(zì)身免疫性疾病發病機制的核心。
B淋巴細胞通過産生保護性抗體在對(duì)抗病原體和(hé)腫瘤的免疫反應中起着至關重要的作(zuò)用(yòng)。它們也(yě)與自(zì)身免疫的發生和(hé)控制有關,其中免疫反應指向自(zì)身抗原。不同的B細胞群表現(xiàn)出保護和(hé)自(zì)我毀滅的能(néng)力。目前了(le)解最多的B淋巴細胞是介導适應性體液反應的傳統骨髓來(lái)源的B2細胞。在包括類風(fēng)濕關節炎(RA)和(hé)系統性紅(hóng)斑狼瘡(SLE)在内的各種自(zì)身免疫性疾病中,B2細胞負責産生自(zì)身抗體、将内源性肽提呈給自(zì)身反應性T細胞以及分泌促炎細胞因子。除了(le)B2細胞外(wài),還有一群獨特的分泌天然抗體的淋巴細胞,稱爲B1細胞。B1細胞産生于胎兒肝髒,在腹膜中發現(xiàn),并具有靜息自(zì)我更新的能(néng)力。B1細胞産生針對(duì)細菌多糖抗原的天然IgM,并表現(xiàn)出顯著的自(zì)我反應性。B1細胞産生的自(zì)身抗體和(hé)細胞因子可介導自(zì)身免疫。然而,B1細胞來(lái)源的自(zì)身抗體也(yě)有助于清除凋亡細胞(自(zì)身抗原的主要來(lái)源),并且似乎可促進腸道(dào)穩态。同時(shí),調節性B細胞(regulatory B cells, Bregs)通過分泌IL-10、TGF -β等免疫調節細胞因子,對(duì)自(zì)身反應性淋巴細胞發揮接觸依賴性抑制作(zuò)用(yòng),在耐受發生中發揮明(míng)确的作(zuò)用(yòng)。
自(zì)身免疫中B細胞功能(néng)障礙的原因尚不完全明(míng)确。雖然代謝(xiè)是細胞活動的明(míng)确調節因素,但(dàn)直到(dào)最近才被認爲是健康和(hé)疾病中B細胞功能(néng)的決定因素。生理(lǐ)上(shàng),代謝(xiè)使B細胞正常發育、分化和(hé)抗體分泌。不出所料,不同發育和(hé)成熟階段的B細胞的不同表型反映在代謝(xiè)活性的顯著差異上(shàng)。此外(wài),不同的B細胞亞群,特别是B1和(hé)B2細胞,利用(yòng)不同的代謝(xiè)途徑。代謝(xiè)是抗自(zì)身免疫檢查點缺失和(hé)無力的重要組成部分,代謝(xiè)失調與逃避耐受性檢查點和(hé)自(zì)身反應性B細胞功能(néng)增強相關。除了(le)傳統的細胞代謝(xiè)外(wài),B細胞還依賴于自(zì)噬,這(zhè)是一種主要對(duì)受損細胞成分進行降解處理(lǐ)的機制。自(zì)噬維持營養剝奪期間的代謝(xiè)穩态,并支持漿細胞(PC)的長期活力。病理(lǐ)學上(shàng),自(zì)噬似乎支持自(zì)身反應性B細胞破壞自(zì)身免疫檢查點,被固有免疫信号激活,并将自(zì)身抗原提呈給T淋巴細胞。
傳統的B2 B細胞的發育必須産生大(dà)量的具有抗體分泌能(néng)力的細胞庫,同時(shí)清除具有自(zì)身反應性抗原特異性的細胞。此外(wài),快(kuài)速生長和(hé)增殖必須在代謝(xiè)困難的骨髓環境中實現(xiàn),需要仔細平衡合成代謝(xiè)和(hé)分解代謝(xiè)信号傳導。前者主要由c-Myc和(hé)線粒體雷帕黴素靶蛋白(bái)複合物(mTORC)信号通路介導,通過上(shàng)調糖酵解和(hé)氧化磷酸化(OXPHOS)來(lái)促進蛋白(bái)質合成和(hé)細胞生長。mTORC信号通路在B細胞發育過程中起着至關重要的作(zuò)用(yòng)。在小(xiǎo)鼠中,敲除mtorc1相關蛋白(bái)Raptor可阻止白(bái)細胞介素(IL) - 7驅動的前b細胞發育。沒有mTORC1信号傳導,前b細胞向前b細胞轉化的能(néng)力較低(dī),而前b細胞是表達前b細胞受體(BCR)的前體,BCR由成熟的免疫球蛋白(bái)重鏈和(hé)替代輕鏈組成。與mTORC1不同,mTORC2在早期B細胞發育中的作(zuò)用(yòng)一直存在争議(yì),盡管它似乎有助于外(wài)周B細胞的成熟。特别是,在B細胞發育的終末階段,mTORC2參與調節mTORC1和(hé)c-Myc的活性。
雖然mTORC信号通路在B細胞發育過程中很(hěn)重要,但(dàn)過度的合成代謝(xiè)活動是有害的。事(shì)實上(shàng),在Fnip1缺失後,B細胞的發育在大(dà)的前B細胞階段受到(dào)損害。Fnip1與5 '腺苷酸活化蛋白(bái)激酶(AMPK)相互作(zuò)用(yòng),AMPK是一種能(néng)量應激傳感器和(hé)分解代謝(xiè)調節因子,與mTORC1相互作(zuò)用(yòng)。雖然在Fnip1缺失的情況下(xià),AMPK可以促進分解代謝(xiè),但(dàn)其抑制mTORC信号傳導的能(néng)力受損。無節制的合成代謝(xiè)導緻前b細胞在前bcr交聯後更易發生凋亡。代謝(xiè)是細胞死亡的調節因子,在抗原刺激下(xià)控制B細胞前體活力,這(zhè)一概念對(duì)對(duì)抗自(zì)身免疫的耐受性檢查點有意義。
大(dà)前B細胞階段(在此期間,pre-BCR在細胞表面表達,并檢測其對(duì)骨髓内自(zì)身抗原的親和(hé)力)代表了(le)自(zì)身免疫檢查點和(hé)代謝(xiè)易感性時(shí)期。盡管其在B細胞發育過程中的必要性存在争議(yì),但(dàn)葡萄糖代謝(xiè)代表了(le)這(zhè)種脆弱性的一個例子。大(dà)前B細胞的快(kuài)速增殖被認爲是通過上(shàng)調的葡萄糖代謝(xiè)來(lái)維持的,因爲大(dà)前B細胞比其他(tā)前體細胞需要更多的葡萄糖。大(dà)的前B細胞經曆了(le)顯著的氧化應激,易受糖酵解抑制,從(cóng)而損害其向小(xiǎo)的前B細胞轉化。通過自(zì)身反應性pre-BCR發出的信号可驅動磷脂酰肌醇3 -激酶(PI3K) -蛋白(bái)激酶B (Akt) -mTORC1通路的過度激活,由此産生的代謝(xiè)應激可誘導負性選擇。非自(zì)身反應性pre-BCR的激活不影響前B或白(bái)血病前B(pre-B ALL)細胞的Akt活化或活力。相反,自(zì)身反應性pre-BCR的激活可誘導akt依賴性快(kuài)速細胞死亡。在pre-B ALL細胞中,PI3K抑制劑磷酸酶和(hé)張力蛋白(bái)同源物(PTEN)的缺失會(huì)增加糖酵解通量,但(dàn)合成代謝(xiè)的增加會(huì)導緻ATP耗竭和(hé)細胞死亡。這(zhè)些(xiē)變化可被mTORC1抑制劑雷帕黴素逆轉,表明(míng)自(zì)身反應性pre-BCR下(xià)遊PI3K-Akt-mTORC1通路的過度激活導緻能(néng)量危機。PTEN在前B細胞的發育中起着至關重要的作(zuò)用(yòng),降低(dī)其對(duì)凋亡的易感性。雖然PTEN缺失影響B細胞前體的非代謝(xiè)特征,如B淋巴轉錄因子的表達,這(zhè)些(xiē)結果進一步強調了(le)平衡代謝(xiè)程序在B細胞發育中的重要性。
除了(le)控制發育的其他(tā)方面外(wài),B淋巴細胞轉錄因子本身還對(duì)B細胞前體細胞施加代謝(xiè)限制,這(zhè)可能(néng)是爲了(le)實現(xiàn)過度激活誘導的細胞死亡。轉錄因子PAX5和(hé)IKZF1的突變在急性淋巴細胞白(bái)血病中常見,提示它們的表達可能(néng)帶來(lái)選擇劣勢。PAX5和(hé)IKZF1在pre-B ALL細胞中的可誘導重建降低(dī)了(le)葡萄糖攝取和(hé)ATP合成,促進了(le)細胞死亡。值得注意的是,無功能(néng)IKZF1的B細胞特異性表達使小(xiǎo)鼠易發生自(zì)身免疫,這(zhè)支持了(le)這(zhè)些(xiē)轉錄因子可能(néng)在B細胞發育過程中發揮耐受作(zuò)用(yòng)的觀點。代謝(xiè)受限也(yě)是B細胞無能(néng)的一個突出特征。除了(le)凋亡和(hé)受體編輯外(wài),耐受發生還可通過誘導無反應性作(zuò)用(yòng)于自(zì)身反應性B細胞及其前體細胞,使B細胞對(duì)抗原刺激反應低(dī)下(xià)。無能(néng)是早期過渡性B細胞從(cóng)骨髓流出後耐受的主要機制。無反應性B細胞的特征是對(duì)BCR或toll樣受體(TLR) 4刺激産生的PI3K信号傳導抑制和(hé)代謝(xiè)重編程受損。據推測,代謝(xiè)抑制可提高(gāo)無反應性自(zì)身反應性B細胞的活化阈值。
鑒于自(zì)噬在促進代謝(xiè)穩态中的作(zuò)用(yòng),科學家們探索了(le)自(zì)噬在B2細胞發育中的作(zuò)用(yòng)。在Rag1 - / -小(xiǎo)鼠的胎肝中,使用(yòng)缺乏關鍵自(zì)噬基因Atg5的細胞進行重建,表明(míng)前b細胞階段的發育受阻。當Atg5缺失局限于成熟B細胞時(shí),脾髒和(hé)淋巴結B細胞群不受影響,這(zhè)意味着自(zì)噬是B2細胞發育所必需的,而不是外(wài)周維持所必需的。然而,使用(yòng)Cd79a-cre進行條理(lǐ)性敲除後,在Atg5表達缺失的情況下(xià)發生前b細胞向前b細胞的轉化。相反,自(zì)噬是維持外(wài)周成熟B細胞群所必需的。考慮到(dào)自(zì)噬抑制劑mTORC1在發育中的關鍵作(zuò)用(yòng),自(zì)噬在B2細胞前體細胞中可能(néng)是可控性的這(zhè)一點可能(néng)并不令人驚訝。
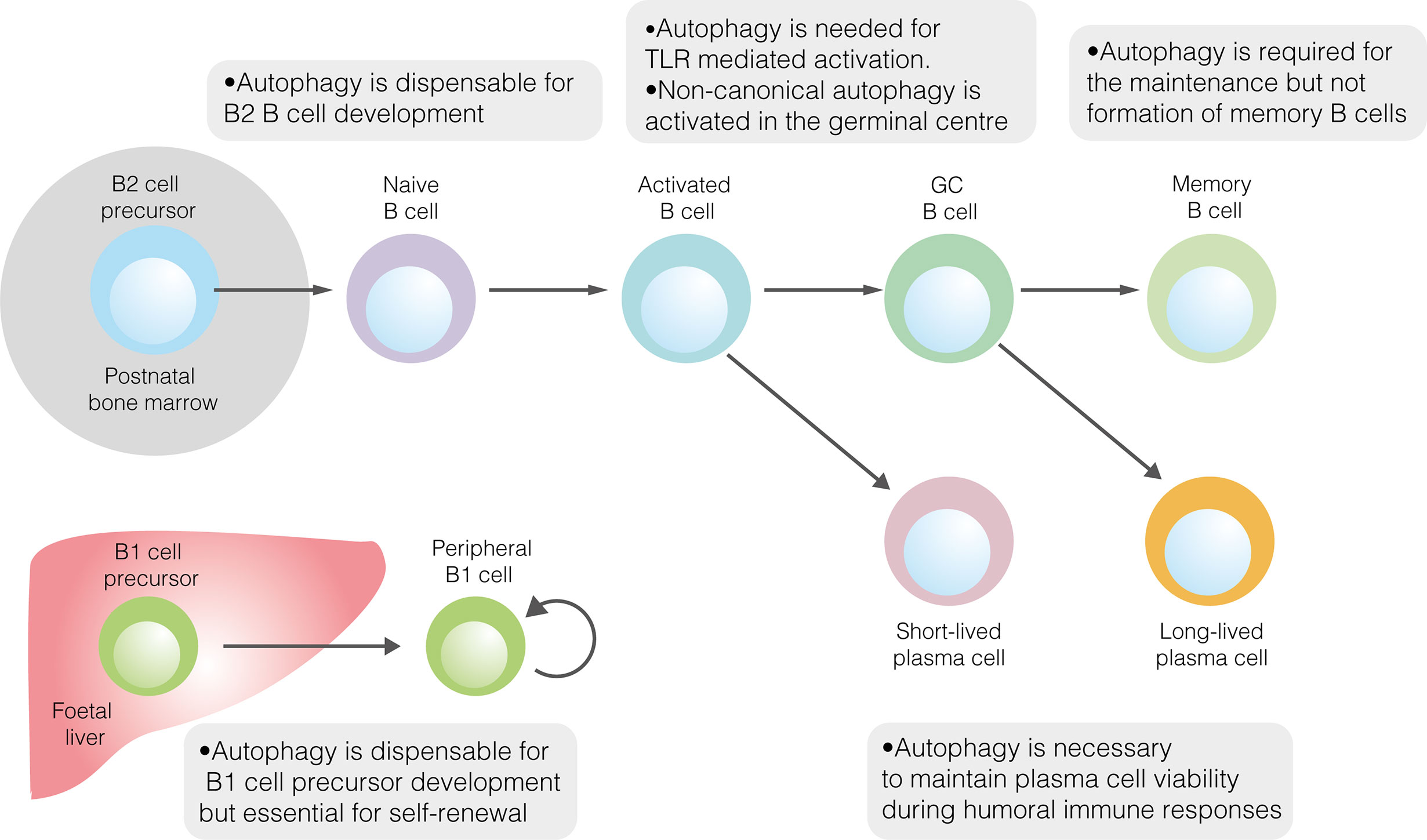
與B2細胞在骨髓中的發育相反,先天樣B1細胞在胎兒肝髒中發育,然後主要遷移到(dào)腹膜和(hé)胸膜。考慮到(dào)B1細胞占據不同的生态位并依靠自(zì)我更新來(lái)維持群體,它們的代謝(xiè)表型與傳統B2細胞的代謝(xiè)表型毫不奇怪。與濾泡B2細胞相比,腹膜B1細胞的特征是更大(dà)的葡萄糖攝取,更高(gāo)的糖酵解率和(hé)OXPHOS,以及對(duì)糖酵解抑制的更高(gāo)敏感性。從(cóng)機制上(shàng)講,葡萄糖代謝(xiè)的升高(gāo)可能(néng)是由高(gāo)水(shuǐ)平的c-Myc表達驅動的。與它們在腹膜富含脂質環境中的定位一緻,B1a細胞廣泛獲取外(wài)源性脂肪酸,同時(shí)也(yě)利用(yòng)内源性脂肪酸合成。
自(zì)噬也(yě)在B1細胞中進行了(le)研究(圖1)。成熟B細胞中自(zì)噬通量的喪失會(huì)導緻B1a細胞的顯著消耗,但(dàn)不會(huì)導緻B1b細胞的顯著消耗。自(zì)噬在B1細胞發育中的具體作(zuò)用(yòng)直到(dào)最近才被探索。在小(xiǎo)鼠中,B1祖細胞群不受自(zì)噬缺失的影響,這(zhè)表明(míng)自(zì)噬不是B1細胞發育所必需的。在沒有自(zì)噬的情況下(xià),小(xiǎo)鼠B1a細胞群的時(shí)間變化支持了(le)這(zhè)一結論。雖然B1a細胞區(qū)室在兩周齡時(shí)是正常的,但(dàn)到(dào)12周齡時(shí),它比野生型小(xiǎo)鼠顯着變小(xiǎo)。總之,這(zhè)些(xiē)結果表明(míng),自(zì)噬是外(wài)周自(zì)我更新所必需的,而不是B1a細胞的分化所必需的。在腹膜B1a細胞中,自(zì)噬似乎對(duì)控制代謝(xiè)基因的表達、脂肪酸的攝取和(hé)脂滴的降解(脂噬)很(hěn)重要。
參考文(wén)獻
Iwan G. A. Raza,B Cell Metabolism and Autophagy in Autoimmunity,Front. Immunol., 07 June 2021,Sec. Autoimmune and Autoinflammatory Disorders,Volume 12 - 2021
獻")








