 聯系我們
聯系我們摘要:
高(gāo)等脊椎動物的蛋白(bái)酪氨酸磷酸酶SHP2(SH2 domain-containing protein-tyrosine phosphatase-2)由ptpn11基因編碼,催化酪氨酸殘基去磷酸化,與其他(tā)能(néng)催化酪氨酸磷酸化的蛋白(bái)酪氨酸激酶共同調節機體内多種信号通路的信号傳導。以往研究表明(míng),SHP2在高(gāo)等脊椎動物T細胞和(hé)B細胞的激活與信号轉導過程中起着重要作(zuò)用(yòng)。
蛋白(bái)酪氨酸磷酸酶(protein tyrosine phosphatases, PTPs)具有催化蛋白(bái)酪氨酸殘基去磷酸化的功能(néng),與能(néng)夠催化蛋白(bái)酪氨酸磷酸化的蛋白(bái)酪氨酸激酶(protein tyrosine kinases, PTKs)作(zuò)用(yòng)相反。在兩者的共同作(zuò)用(yòng)下(xià),蛋白(bái)酪氨酸殘基發生可逆磷酸化,從(cóng)而可以調節細胞膜酶聯受體的信号轉導,對(duì)生物的生長發育具有重要意義。研究表明(míng)PTKs主要與緻癌和(hé)促腫瘤活性相關,而PTPs通常在各種人類腫瘤細胞中發生突變或異常活化,表明(míng)其正常功能(néng)的維持會(huì)對(duì)腫瘤的發展起一定的抑制作(zuò)用(yòng)。蛋白(bái)酪氨酸磷酸酶SHP2是PTP家族中的一員,也(yě)是第一個被證實的原癌基因,廣泛表達于各種組織中,參與多種生物學信号的轉導,是脊椎動物發育中不可缺少的胞内非受體型PTP。人類SHP2蛋白(bái)由ptpn11基因編碼,其氨基酸序列具有一個N端Src同源2 (N-terminal Src homology 2, N-SH2)結構域、一個與N-SH2結構域相鄰的C-SH2結構域和(hé)一個跨度比較大(dà)的PTP結構域。在SHP2分子的C端有兩個磷酸化位點(Y542與Y580)和(hé)一個富含脯氨酸基序的尾部。N-SH2結構域是一種構象開(kāi)關,在正常生理(lǐ)條件下(xià),SHP2的N-SH2結構域與其自(zì)身的PTP結構域相結合,抑制其磷酸酶活性。然而,SHP2如果通過其SH2結構域與酪氨酸磷酸化生長因子受體或接頭蛋白(bái)結合則會(huì)導緻這(zhè)種分子内相互作(zuò)用(yòng)被破壞,使其PTP結構域上(shàng)的催化活性中心暴露,從(cóng)而激活其磷酸酶活性。通過串聯的SH2結構域識别雙磷酸化的配體是該開(kāi)關的組成部分;而C端SH2結構域雖然有助于提高(gāo)結合能(néng)和(hé)特異性,但(dàn)它被激活後并不能(néng)直接起作(zuò)用(yòng)。
SHP2是一種分布廣泛的酪氨酸磷酸酶,作(zuò)爲轉導因子參與細胞内由各種生長因子、激素或細胞因子介導的重要的信号通路。如SHP2可以促進小(xiǎo)GTP結合蛋白(bái)(Rat sarcoma, Ras)/絲裂原活化蛋白(bái)激酶(mitogen-activated protein kinase, MAPK)途徑激活,而Ras/MAPK通路對(duì)細胞的增殖、分化和(hé)凋亡起着重要作(zuò)用(yòng)。當細胞受到(dào)生長因子的刺激後,SHP2作(zuò)爲正調控因子促進受體酪氨酸激酶激活的下(xià)遊細胞外(wài)調節蛋白(bái)激酶(extracellular regulated protein kinases, Erk)/MAPK信号通路,也(yě)可以增強由細胞因子IL-21激活的細胞增殖過程中ERK1/2信号傳導活性。體外(wài)實驗證明(míng),SHP2也(yě)能(néng)夠通過正向調控絲/蘇氨酸蛋白(bái)激酶Akt (serine/threonine protein kinases Akt, Akt)和(hé)Erk1/2信号傳導促進少突膠質細胞的成熟。SHP2在導緻Erk激活的各種促有絲分裂信号傳導途徑中起着正調控作(zuò)用(yòng),但(dàn)也(yě)有研究發現(xiàn)SHP2在介導細胞外(wài)信号調節和(hé)下(xià)遊c-Jun氨基端激酶(c-Jun NH2-terminal kinases, JNK)信号通路的激活方面具有負調控作(zuò)用(yòng)。這(zhè)也(yě)是酪氨酸磷酸酶SHP2在介導Erk和(hé)JNK、MAPK活化中具有負調控作(zuò)用(yòng)的第一個證據。另外(wài),SHP2在高(gāo)等動物适應性免疫系統的免疫應答(dá)反應中也(yě)發揮着重要作(zuò)用(yòng)。Hoff等發現(xiàn)SHP-2在T細胞受體(T cell receptor, TCR)介導的免疫反應中起重要作(zuò)用(yòng),促進炎症細胞因子的分泌、趨化因子誘導的遷移以及活化細胞的凋亡。
七鰓鳗(Petromyzoniformes)作(zuò)爲目前世界上(shàng)僅存的最原始無颌類脊椎動物之一,承接了(le)從(cóng)無脊椎動物到(dào)脊椎動物的進化曆程,被稱爲“活化石”。由于其在進化上(shàng)的特殊地位,七鰓鳗成爲研究脊椎動物起源與進化的關鍵物種。研究證明(míng),在七鰓鳗體内不僅存在着先天性免疫,同時(shí)存在着适應性免疫,而七鰓鳗的适應性免疫不同于高(gāo)等脊椎動物。高(gāo)等脊椎動物的适應性免疫依賴于免疫球蛋白(bái)(Ig)及TCR和(hé)B細胞受體(BCR)分子介導,七鰓鳗的适應性免疫系統則是由可變淋巴受體(variable lymphocyte receptors, VLR)介導。在無颌類脊椎動物中,其适應性免疫系統中最顯著的特征是使用(yòng)富含亮(liàng)氨酸的重複模塊來(lái)組裝可變淋巴細胞受體基因(VLRA、VLR B和(hé)VLRC)。爲了(le)更加清晰地了(le)解無颌類脊椎動物是否存在作(zuò)爲适應性免疫應答(dá)信号傳導的重要調控因子SHP2,本研究在日本七鰓鳗(Lampetra japonica)中克隆并鑒定了(le)與高(gāo)等脊椎動物SHP2同源的分子,研究SHP2在日本七鰓鳗免疫應答(dá)過程中的作(zuò)用(yòng),揭示SHP2家族的系統發生,爲探索高(gāo)等脊椎動物适應性免疫系統的早期發生及其進化曆程提供一定的線索。
來(lái)自(zì)遼甯師範大(dà)學生命科學學院、大(dà)連工(gōng)業大(dà)學的研究人員通過PCR 擴增獲取其Lja-SHP2開(kāi)放(fàng)閱讀框序列,并構建到(dào)原核表達載體 pET-32a中,成功在大(dà)腸杆菌中實現(xiàn)重組蛋白(bái)表達并制備了(le)其兔源多克隆抗體。用(yòng)混合菌免疫刺激日本七鰓鳗後,通過實時(shí)熒光定量PCR和(hé)免疫印迹方法檢測了(le)Lja-SHP2在日本七鰓鳗免疫相關組織中mRNA和(hé)蛋白(bái)水(shuǐ)平表達譜。結果顯示,混合菌免疫刺激後,Lja-SHP2 mRNA和(hé)蛋白(bái)表達在外(wài)周血白(bái)細胞和(hé)髓樣小(xiǎo)體中無顯著變化,而在鰓組織中顯著性上(shàng)調(P<0.05),說明(míng)Lja-SHP2在混合菌刺激後主要參與了(le)鰓組織的免疫應答(dá)反應。爲了(le)進一步探究Lja-SHP2與淋巴細胞亞群免疫應答(dá)反應的相關性,本研究分别使用(yòng)B細胞有絲分裂原脂多糖(lipopolysaccharide, LPS)和(hé)T細胞的有絲分裂原植物凝集素(phytohemagglutinin, PHA)免疫刺激日本七鰓鳗。經LPS免疫刺激後,與對(duì)照組相比,白(bái)細胞中Lja-SHP2蛋白(bái)表達顯著上(shàng)調,鰓組織和(hé)髓樣小(xiǎo)體沒有顯著性差異表達;但(dàn)經PHA免疫刺激後,與對(duì)照組相比,白(bái)細胞、鰓組織和(hé)髓樣小(xiǎo)體3種組織中Lja-SHP2均有上(shàng)調,尤其在白(bái)細胞中上(shàng)調最爲顯著,大(dà)約是對(duì)照組的2.5倍,說明(míng)Lja-SHP2參與了(le)日本七鰓鳗由PHA介導的免疫應答(dá)反應。由于PHA能(néng)刺激日本七鰓鳗鰓組織中VLRA +淋巴細胞的活化,這(zhè)表明(míng)Lja-SHP2可能(néng)參與了(le)PHA介導的VLRA +淋巴細胞亞群的免疫應答(dá)反應。
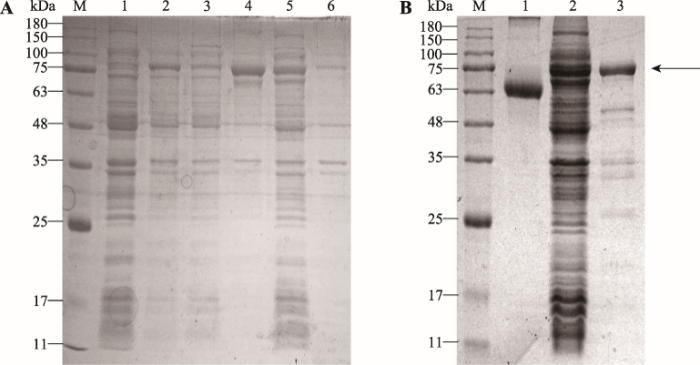
SDS-PAGE檢測Lja-SHP2的重組表達、可溶性及純化結果
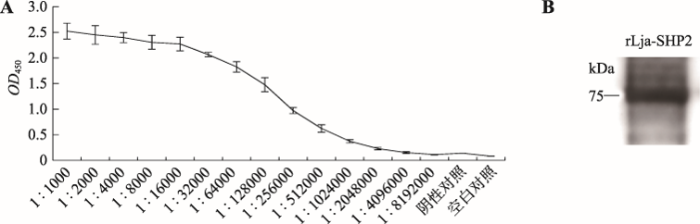
Lja-SHP2多克隆抗體效價與特異性檢測
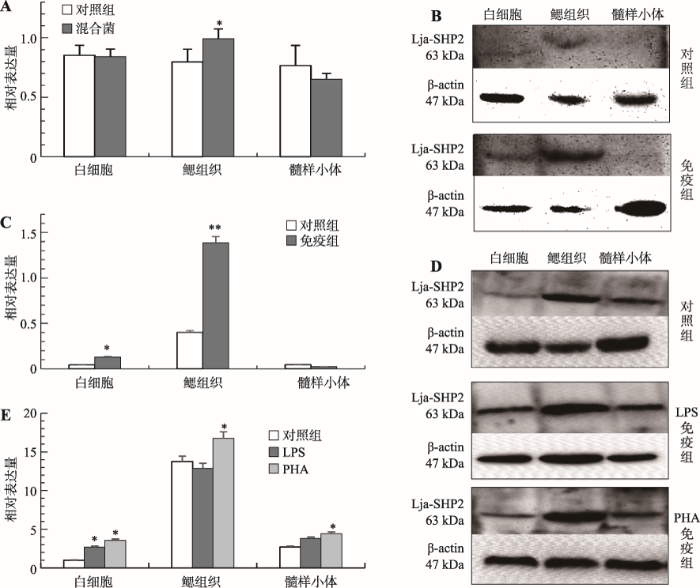
Lja-SHP2 mRNA和(hé)蛋白(bái)質在各免疫相關組織中的相對(duì)表達量
SHP2作(zuò)爲蛋白(bái)酪氨酸磷酸酶家族成員之一,參與多條信号通路,如RAS/MAPK通路、PI3K/AKT通路及JAK/STAT通路等,不僅爲脊椎動物胚胎發育所必需,而且也(yě)參與多種組織特異性細胞的增殖、分化和(hé)凋亡的調控。SHP2也(yě)是一種特殊的蛋白(bái)酪氨酸磷酸酶,它既可以作(zuò)爲正調節因子調節下(xià)遊信号轉導通路,也(yě)可以在特定條件下(xià)發揮負調控作(zuò)用(yòng)。研究發現(xiàn),與正常T細胞相比,在缺失ptpn11的T細胞中Erk激酶活化受到(dào)阻礙;表明(míng)SHP2通過調節Erk途徑促進TCR信号傳導,是pre-TCR和(hé)TCR促進T細胞成熟和(hé)增殖的常用(yòng)信号傳感器。另外(wài),SHP2通過介導T細胞發育和(hé)功能(néng)參與多種信号傳導途徑,可以調節細胞因子依賴性粒細胞生成,也(yě)是嗜酸性粒細胞分化的關鍵調節因子。
獻")








